AUCTORES
Globalize your Research

Research Article | DOI: https://doi.org/10.31579/2766-2314/038
*Corresponding Author: Azab Elsayed Azab, Department of Physiology, Faculty of Medicine, Sabratha University, Sabratha Libya.
Citation: Marwan T. M. Abofila, Azab E. Azab and Asma N. Bshena (2021) Skeletal Muscles: Insight into Embryonic Development, Satellite Cells, Histology, Ultrastructure, Innervation, Contraction and Relaxation, Causes, Pathophysiology, and Treatment of Volumetric Muscle Injury. J, Biotech. and Bioprocessing 2(4); DOI: 10.31579/2766-2314/038
Copyright: © 2021, Marwan T. M. Abofila, This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 03 April 2021 | Accepted: 19 April 2021 | Published: 23 April 2021
Keywords: embryonic development of skeletal muscles, histological and ultrastructure, innervation, contraction and relaxation, volumetric muscle injury
Background: Skeletal muscles are attached to bone and are responsible for the axial and appendicular movement of the skeleton and for maintenance of body position and posture.
Objectives: The present review aimed to high light on embryonic development of skeletal muscles, histological and ultrastructure, innervation, contraction and relaxation, causes, pathophysiology, and treatment of volumetric muscle injury. The heterogeneity of the muscle fibers is the base of the flexibility which allows the same muscle to be used for various tasks from continuous low-intensity activity, to repeated submaximal contractions, and to fast and strong maximal contractions. The formation of skeletal muscle begins during the fourth week of embryonic development as specialized mesodermal cells, termed myoblasts. As growth of the muscle fibers continues, aggregation into bundles occurs, and by birth, myoblast activity has ceased. Satellite cells (SCs), have single nuclei and act as regenerative cells. Satellite cells are the resident stem cells of skeletal muscle; they are considered to be self-renewing and serve to generate a population of differentiation-competent myoblasts that will participate as needed in muscle growth, repair, and regeneration. Based on various structural and functional characteristics, skeletal muscle fibres are classified into three types: Type I fibres, Type II-B fibres, and type II-A fibres. Skeletal muscle fibres vary in colour depending on their content of myoglobin. Each myofibril exhibits a repeating pattern of cross-striations which is a product of the highly ordered arrangement of the contractile proteins within it. The parallel myofibrils are arranged with their cross-striations in the register, giving rise to the regular striations seen with light microscopy in longitudinal sections of skeletal muscle. Each skeletal muscle receives at least two types of nerve fibers: motor and sensory. Striated muscles and myotendinous junctions contain sensory receptors that are encapsulated proprioceptors. The process of contraction, usually triggered by neural impulses, obeys the all-or-none law. During muscle contraction, the thin filaments slide past the thick filaments, as proposed by Huxley's sliding filament theory. In response to a muscle injury, SCs are activated and start to proliferate; at this stage, they are often referred to as either myogenic precursor cells (MPC) or myoblasts. In vitro, evidence has been presented that satellite cells can be pushed towards the adipogenic and osteogenic lineages, but contamination of such cultures from non-myogenic cells is sometimes hard to dismiss as the underlying cause of this observed multipotency. There are, however, other populations of progenitors isolated from skeletal muscle, including endothelial cells and muscle-derived stem cells (MDSCs), blood-vessel-associated mesoangioblasts, muscle side-population cells, CD133+ve cells, myoendothelial cells, and pericytes. Volumetric muscle loss (VML) is defined as the traumatic or surgical loss of skeletal muscle with resultant functional impairment. It represents a challenging clinical problem for both military and civilian medicine. VML results in severe cosmetic deformities and debilitating functional loss. In response to damage, skeletal muscle goes through a well-defined series of events including; degeneration (1 to 3days), inflammation, and regeneration (3 to 4 weeks), fibrosis, and extracellular matrix remodeling (3 to 6 months).. Mammalian skeletal muscle has an impressive ability to regenerate itself in response to injury. During muscle tissue repair following damage, the degree of damage and the interactions between muscle and the infiltrating inflammatory cells appear to affect the successful outcome of the muscle repair process. The transplantation of stem cells into aberrant or injured tissue has long been a central goal of regenerative medicine and tissue engineering.
Conclusion: It can be concluded that the formation of skeletal muscle begins during the fourth week of embryonic development as specialized mesodermal cells, termed myoblasts, by birth myoblast activity has ceased. Satellite cells are considered to be self-renewing, and serve to generate a population of differentiation-competent myoblasts. Skeletal muscle fibres are classified into three types. The process of contraction, usually triggered by neural impulses, obeys the all-or-none law. VML results in severe cosmetic deformities and debilitating functional loss. Mammalian skeletal muscle has an impressive ability to regenerate itself in response to injury. The transplantation of stem cells into aberrant or injured tissue has long been a central goal of regenerative medicine and tissue engineering.
Skeletal muscles are attached to bone and are responsible for the axial and appendicular movement of the skeleton and maintenance of body position and posture. Also, skeletal muscles of the eye (extra-ocular muscles) provide precise eye movement [1]. Based on various structural and functional characteristics, skeletal muscle fibres are classified into three types: Type I fibres, Type II-B fibres, and type II-A fibres [2].
Mammalian skeletal muscles are heterogeneous. The functional unit of the motor system, the motor unit, is composed of a motor neuron and a bunch of muscle fibers with similar, but not identical, structural and functional properties. The heterogeneity of the muscle fibers is the base of the flexibility which allows the same muscle to be used for various tasks from continuous low-intensity activity (e.g., posture), to repeated submaximal contractions (for example, locomotion), and to fast and strong maximal contractions (jumping, kicking) [2]. Part of the myoblast population does not fuse and differentiate but remains as a group of mesenchymal cells called muscle satellite cells located on the external surface of muscle fibers inside the developing external lamina. Satellite cells proliferate and produce new muscle fibers following muscle injury [3].
Muscle contraction obeys the "all-or-none law" and is followed by muscle relaxation. Contraction effectively reduces the resting length of the muscle fiber by an amount that is equal to the sum of all shortenings that occur in all sarcomeres of that particular muscle cell [4].
The present review aimed to high light on embryonic development of skeletal muscles, histological and ultrastructure, innervation, contraction and relaxation, causes, pathophysiology, and treatment of volumetric muscle injury.
The formation of skeletal muscle begins during the fourth week of embryonic development as specialized mesodermal cells, termed myoblasts, which begin rapid mitotic division. The cytoplasmic fusion of myoblasts forms what is known as myotubes. By week nine of development, these can be identified as multinucleate skeletal muscle cells, termed muscle fibers (Figure .1) [5]. By the fifth month, the muscle fibers start accumulating protein filaments important in muscle contraction. As growth of the muscle fibers continues, aggregation into bundles occurs, and by birth, myoblast activity has ceased [6].
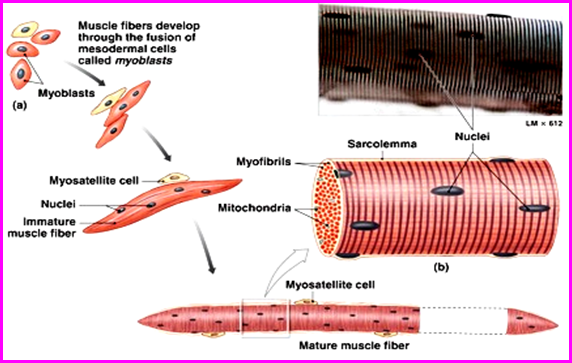
Satellite cells (SCs), have single nuclei and act as regenerative cells. They are located in shallow depressions on the muscle cell's surface, sharing the muscle fiber's external lamina. The chromatin network of the satellite cell nucleus is denser and coarser than that of the muscle fiber [4]. Satellite cells are the resident stem cells of skeletal muscle; they are considered to be self-renewing and serve to generate a population of differentiation-competent myoblasts that will participate as needed in muscle growth, repair and regeneration [7].
Satellite cells were first described by Mauro [8] and are so called due to its peripheral location on the skeletal muscle myofiber, where it lies between the sarcolemma of the myofiber cell and its surrounding basal lamina, they have unique morphological characteristics, including abundant cytoplasm, a small nucleus with increased amounts of heterochromatin and reduced organelle content. These features reflect the fact that satellite cells are mitotically quiescent and transcriptionally less active than myonuclei [9].
Satellite cells (SCs) have been shown to express different types of genetic markers, about 10% of them are (Pax7+/Myf5−) stem cells, and 90% are committed myogenic progenitors (Pax7+/Myf5+) [10]. In mature muscle tissue, satellite cells occur as a small, dispersed population of mitotically and physiologically quiescent cells, marked by their expression of cell-surface markers, including syndecan-4 [11] and α7 integrin [12]. More recently, variation in the expression of various non-specific myogenic markers, in satellite cells, such as nestin [13], b1-integrin [14], and Syndecan-4 [15] have also been described. Despite the evident heterogeneity, the phenotypical characteristics of these subpopulations are hard to elucidate because their behavior in vitro has been difficult to investigate.
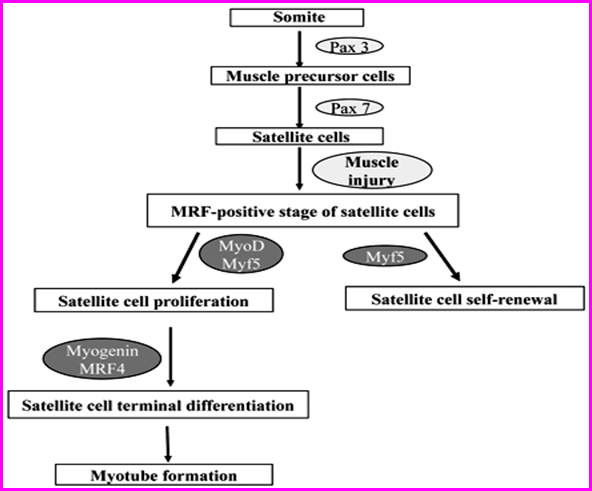
Satellite cells originate from somites [7, 16]. SCs are present in healthy adult mammalian muscle as quiescent cells and represent 2.5%–6% of all nuclei of a given muscle fiber. Quiescent SCs express characteristic (although not unique) markers. In the mouse, the most widely used of these markers is the transcription factor paired box 7 (Pax7) [17], which is essential for SC specification and survival [18]. In contrast, Pax3 is expressed only in quiescent SCs in a few specific muscle groups (Figure. 2) [19]. In humans, markers of both quiescent and activated SCs do not fully correspond to those in the mouse, and relatively little is known about them due to the difficulty of obtaining human tissue. For example, although CD34 is a marker of SCs in mice, it does not mark SCs in human muscle [20].
Skeletal muscles are sheathed by a tough layer of connective tissue called the epimysium. The epimysium anchors muscle tissue to tendons at each end, where the epimysium becomes thicker and collagenous. It also protects muscles from friction against other muscles and bones. Within the epimysium are multiple bundles called fascicles, each of which contains 10 to 100 or more muscle fibers collectively sheathed by perimysium. Besides surrounding each fascicle, the perimysium is a pathway for nerves and the flow of blood within the muscle. The thread-like muscle fibers are the individual muscle cells (myofibers), and each cell is encased within its endomysium which is composed of reticular fibers and an external lamina (basal lamina) [21].
One of the most important roles of this connective tissue is to transmit the mechanical forces generated by the contracting muscle cell/fibers because individual muscle cells seldom extend from one end of a muscle to the other [3].
The muscle penetrates by blood vessels within the septa of connective tissue that made endomysium have a rich capillary network. Lymphatic vessels and larger blood vessels are found in the other connective tissue layers [3].
The cell contractile apparatus largely occupied both sarcoplasm & the cytoplasm of each myofiber. This is made up of myofibrils arranged in sarcomeres, which are the cell contractile units. The myofibrils are longitudinally arranged cylindrical structures, each 1 to 2μm in diameter [21].
Skeletal muscle fibres vary in colour (red, white, and intermediate) depending on their content of myoglobin (myoglobin stores oxygen until needed by the mitochondria). Fibers of skeletal muscle contract at different rates, based on their ability to partition adenosine triphosphate (ATP). Furthermore, fibers of skeletal muscle vary regarding the processes of metabolic that they use to generate ATP. It also differs in terms of the fatigue onset. Based on various structural and functional characteristics, skeletal muscle fibres are classified into three types: Type I fibres, Type II B fibres and type II A fibres [2].
Fibers of Type I: These fibers, also known as fibers of slow-twitch or slow-oxidizing, contain myoglobin with large amounts, several mitochondria, and several blood capillaries. Fibers of type I are red, which are slow regarding the rate of split ATP, and contraction velocity, but have a high ability to generate ATP by processes of oxidative metabolic, and very resistant to fatigue. Such fibres are found in large numbers in the postural muscles of the neck [22].
Fibers of Type II-A: These fibers, also known as fibers of fast-twitch or fast-oxidizing, contain myoglobin with large amounts, several mitochondria, and several blood capillaries. Fibres of type II-A are red and resistant to fatigue which are very fast with concerning the rate of split ATP, as well as have a very high ability to generate ATP by processes of oxidative metabolic and have a rapid contraction. Such fibres are infrequently found in humans [22].
Fibers of Type II-B: These fibers, also known as fibers of fast-twitch or fast- glycolytic, contain myoglobin with low amounts but contain a large content of glycogen, and relatively few of both mitochondria and blood capillaries. Fibres of type II-B are white and easy to fatigue, geared to generate ATP by processes of anaerobic metabolic, not able to provide fibres of skeletal muscle constantly with enough ATP, split ATP at a rapid rate and have a rapid contraction rate. Such fibres are found in large numbers in the muscles of the arms [22].
In different classes of vertebrates, Fiber of muscle type diversification is observed since early stages of development. The notion of Satellite cell heterogeneity is supported by the finding that satellite cells from adult mammalian muscle tend to form fibers with a phenotype reflecting their fiber type of origin [23].
The finding that regenerating fast and slow skeletal muscle show a different myosin heavy chain (MyHC) composition when stimulated with the same impulse pattern in the absence of innervation, probably reflecting intrinsically different satellite cells [24].
The very orderly arrangement of contractile proteins within each myofibril, lead to exhibits a frequent pattern of cross-striations. This can only be seen with electron microscopy. Furthermore, the parallel myofibrils are arranged with their cross-striations in the register, giving rise to the regular striations seen with light microscopy in longitudinal sections of skeletal muscle [21].
The dark bands are known as A bands (anisotropic with polarized light) and the light bands as I bands (isotropic with polarized light). Both A and I bands are divided by narrow regions of varying intensities. A dense line, the Z line, also called the Z disc, bisects the light I band. Also the dark band A is split into a less intense or light region called the H band. In addition, the division of the light band H is a narrow and dense line known as the M line. The M line is best demonstrated in electron micrographs, although in ideal H&E preparations it can be detected in the Light microscope [1].
The pattern of A and I bands in sarcomeres is mainly due to the regular arrangement of two types of myofilaments - thin and thick - which lie parallel to the long axis of the myofibrils in a symmetrical pattern. [3].
The thick filaments occupying the A band at the center of the sarcomere are 1.6µm long and 15 nm wide. The thin filaments have one end attached to the Z line and these filaments run between and parallel to the thick filaments. Thin filaments are 8 nm wide and 1.0m long. This arrangement makes the I band consist of part of the thin filaments that do not overlap the thick filaments, which stained lighter staining. A-bands consist mainly of thick filaments as well as interlacing portions of thin filaments. The A band shows the central light zone, the H zone, that corresponds to a region consisting only of the rod-like portions of the myosin molecule with no thin filaments present. Bisecting the H zone is the M line, a region where lateral connections are made between adjacent thick filaments. The main proteins present in the M lineage region are myomesin, a myosin-binding protein that holds thick filaments in place, and creatine kinase, which catalyzes the transfer of phosphate groups from phosphocreatine (a storage form of high-energy phosphate groups) to adenosine diphosphate (ADP), thus helping to supply adenosine triphosphate (ATP) for muscle contraction [25].
Thick and thin filaments overlap for some distance in band A. As a consequence, a cross section in the region of filament overlap shows each thick filament surrounded by six thin filaments in the form of a hexagon [3].
Thin filaments are composed of F-actin, associated with tropomyosin, which also forms a long fine polymer, and troponin, a globular complex of three subunits. Thick filaments consist primarily of myosin. Myosin and actin together represent 55% of the total protein of striated muscle.
F-actin consists of long filamentous polymers containing two strands of globular (G-actin) monomers, 5.6 nm in diameter, twisted around each other in a double helical formation. G-actin molecules are asymmetric and polymerize to produce a filament with polarity (Figure. 3). Each G-actin monomer contains a binding site for myosin. Actin filaments, which are anchored perpendicularly on the Z line by the actin-binding protein -actinin, exhibit opposite polarity on each side of the line [3].

Each tropomyosin sub0unit is a long, thin molecule about 40 nm in length containing two polypeptide chains, which assembles to form a long polymer located in the groove between the two twisted actin strands [3].
Troponin is a complex of three subunits: TnT, which attaches to tropomyosin; TnC, which binds calcium ions; and TnI, which inhibits the actin-myosin interaction. Troponin complexes are attached at specific sites at regular intervals along each tropomyosin molecule [3].
Thick filaments are composed of myosin II molecules aligned end to end. Every thick filament consists of 200 to 300 myosin II molecules. Each myosin II molecule (150 nm in length; 2 to 3 nm in diameter) is composed of two identical heavy chains and two pairs of light chains. The heavy chains (two globular heads and two rod-like polypeptide chains) are wrapped around each other in an α-helix. The heavy chains can be cleaved by trypsin into:
Light meromyosin, a rod-like tail composed of the two rod-like polypeptide chains wrapped around each other.
Heavy meromyosin, two globular heads (resembling golf clubs) with short proximal portions of the two rod-like polypeptide chains wrapped around each other [26].
Light meromyosin works in the appropriate assembly of molecules in the thick bipolar filament. Heavy meromyosin is cleaved by papain into two globular (S1) segments; binds adenosine triphosphate (ATP) and functions in the formation of cross-bridges between the thick and thin myofilaments molecule; and a short, helical, rod-like segment (S2). Light chains are of two types, and one of each type is associated with each S1subfragment of the myosin II molecule. For each heavy chain, therefore, there are two light chains. A myosin II molecule is composed of two heavy chains and four light chains [26].
Each molecule of myosin seems to contain two elastic regions, one at the junction of sub-fragments for S1 and S2 and the other at the junction of light and heavy meromyosin. The elastic region between both light and heavy meromyosins allows each molecule of myosin II to come into contact with the fine filament, forming a cross-bridge between the two types of filaments. And, the flexible region between S1 and S2 subfragments enables the myosin II molecule to drag the thin filament towards the middle of the sarcomere [26].
Largely five proteins maintain the structural organization of myofibrils: Titin, α-Actinin, Cap Z, Nebulin, and Tropomodulin proteins.
Thick filaments are positioned precisely within the sarcomere with the assistance of titin, a large, linear, elastic protein. Each, a thick filament half extend two titin molecules to the adjacent Z disc; thus, four molecules of titin anchor a thick filament between the two Z discs of each sarcomere.
Thin filaments are held in the register by the rod-shaped protein α-actinin, a component of the Z disk that can bind thin filaments in parallel arrays. The excess end of the thin filament is held in place by a protein known as Cap Z, which also prevents the further, or subtraction of G-actin molecules to or from the thin filament, thus helping to maintain its delicate length. Furthermore, nebulin two molecules, a long inelastic protein, are coiled along the entire length of each thin filament, anchoring it further in the Z disk and ensuring that the specific array of thin filaments is maintained. Moreover, nebulin acts as a "ruler," ensuring the precise length of the thin filament. It is assisted in this function by the protein tropomodulin, a cap on the minus end of the thin filament that, similarly to Cap Z, prevents the addition or the deletion of G-actin molecules to or from the thin filament [4].
In skeletal muscle, the smooth endoplasmic reticulum (SER) is specialized for Ca2+ ion sequestration. The depolarization of this membrane of the sarcoplasmic reticulum that outcomes in the release of Ca2+ ions. It begins at the specialized neuromuscular junction on the muscle cell surface. Surface-initiated depolarization signals should diffuse throughout the cell to produce a release of Ca2 + from the internal cisterns of the sarcoplasmic reticulum. In large cells of muscle , such diffusion of the depolarization signal would lead to a contraction wave with contracting of peripheral myofibrils before those more centrally positioned. To ensure uniform contraction, fibers of skeletal muscle have a system of transverse tubules (T). These finger-like invaginations of the sarcolemma form a complex network of tubules that encircles every myofibril near the A-I band boundaries of each sarcomere [3].
At least two types of nerve fibers receives by every skeletal muscle: sensory and motor. The motor nerve acts to stimulate contraction, while the sensory fibers are transmitted to the spindles of the muscles. Besides that, the autonomic fibers provide the vascular elements of skeletal muscle. The motor innervation specificity is the innervated muscle function. If the muscle works carefully, as some muscles in the eye do, a single motor neuron can be responsible for 5-10 fibers of skeletal muscle, while a muscle in the abdominal wall can have up to 1000 fibers under control of a single motor neuron. Each motor neuron forms muscle fibers that are controlled by a motor unit. The muscle fibers of a motor unit contract in unison and follow the all-or-none law of muscle contraction [4].
The myelinated motor nerves branch out inside the peripheral connective tissue, with each nerve producing several terminal branches. At the innervation site, the axon loses its myelin sheath and forms an expanding end located within a trough on the surface of the muscle cells. This structure is known as the myoneural junction or the motor endplate. Only the axon at this site is covered by a thin visceral extension from the cell of Schwann. Within the axon terminal, there are many synaptic vesicles and mitochondria, the previous one containing the acetylcholine of neurotransmitter. There is a space between the axon and muscle, the synaptic cleft, in which there is an amorphous basal lamina matrix from fibers of muscle. The sarcolemma at the junction is thrown into many deep junction folds, which provide a larger surface area. Below the folds of the sarcoplasm lie many nuclei and numerous mitochondria, glycogen granules, and ribosomes [3].
Myotendinous junctions and striated muscles comprise sensory receptors which are encapsulated proprioceptors (L. proprius, one's own, + capio, to take). Muscle spindles are stretch detectors that from among the muscle fascicles, which contain a capsule of connective tissue surrounding a fluid-filled space that consists of a few thin, fibers of nonstriated muscle densely filled with nuclei, and known as intrafusal fibers. Numerous axons of sensory nerves penetrate each muscle spindle and wrap around individual intrafusal fibers. Length changes in (usually stretched) of striated (extrafusal) fibers caused by body movements are detected by sensory nerves and muscle spindles transmit this information to the spinal cord. Mediate reactions of varying complexity by various types of internal and sensory fibers to maintain posture and regulate the activity of opposing muscle groups contributory in motor activities for example walking [3].
In tendons, near the insertion sites of muscle fibers, a connective tissue sheath encapsulates the large collagen bundles of the myotendinous junction. Sensory nerves penetrate this capsule and form another sensory receptor known as (Golgi) tendon organs. Tendon organs detect changes in tension within tendons produced by muscle contraction and act to inhibit motor nerve activity if tension becomes excessive. Because both of these sensory receptors detect increases in tension, they help to regulate the amount of effort required to perform movements that call for variable amounts of muscular force [3].
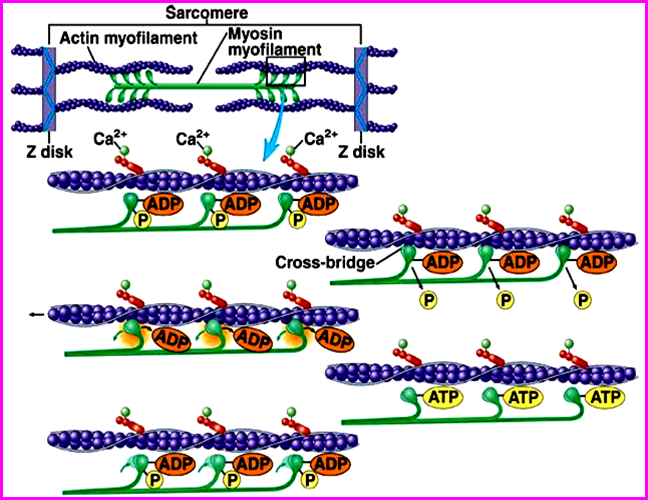
The process of contraction, usually triggered by neural impulses, obeys the all-or-none law, in that a single muscle fiber either contracts as a result of stimulation or does not respond at all. The strength of contraction of a gross anatomical muscle, such as the biceps, is a function of the number of muscle fibers that undergo contraction. The stimulus is transferred at the neuromuscular junction. During muscle contraction, the thin filaments slide past the thick filaments, as proposed by Huxley's sliding filament theory [4].
During contraction, the I band decreases in size as thin filamentspenetrate the A band. The H band—the part of the A band with only thick filaments—diminishes in width as the thin filaments completely overlap the thick filaments. A net result is that each sarcomere, and consequently the whole cell (fiber), is greatly shortened. The following sequence of events leads to contraction in skeletal muscle:
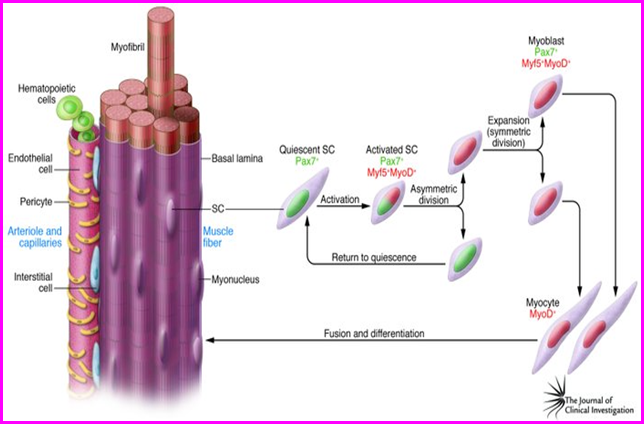
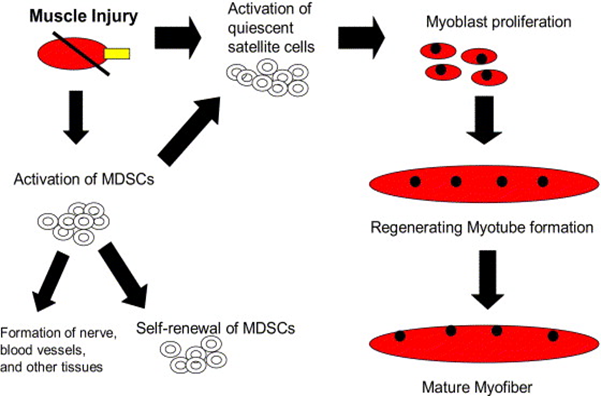
In response to a muscle injury, Satellite cells (SCs) are activated and start to proliferate; at this stage, they are often referred to as either myogenic precursor cells (MPC) or myoblasts [27, 28]. Several signals, deriving both from damaged fibers and infiltrating cells, are involved in SC activation, including; Hepatocyte growth factor HGF [29], Fibroblast growth factor FGF [7], Insulin-like growth factor IGF [30] and Nitric oxide NO [31].
Using a new experimental maneuver that permits clear and correct isolation of SCs from the fiber of origin, it was reported for the first time, that two subpopulations of SCs coexist in fixed proportions on the single fiber: the low proliferative (LPC) and the high proliferative clones (HPC) which show alternative myogenic potential in vitro and retained also in vivo [32].
In adults, grafting an isolated myofiber with a small number of associated satellite cells [33], or even just a single fluorescence-activated cell sorting (FACS)-isolated satellite cell [34], produces many more donor-derived satellite cells in the host muscle than originally transplanted. Such amplification requires extensive proliferation from the donor satellite cells and/or their progeny, showing that self-renewal had occurred. Importantly, donor satellite cells remain viable, being able to again participate in regeneration if the muscle is damaged [33]. Donor-derived satellite cells can also be recovered from muscles after transplantation, and serially transplanted/recovered several more times, showing the extent of their potential for self-renewal [35]. Thus, as satellite cells not only generate differentiated progeny, but also maintain their own population by self-renewal, they can be classified as myogenic stem cells.
The term ‘stem cell’, however, also evokes thoughts of multi-potency. In vitro, evidence has been presented that satellite cells can be pushed towards the adipogenic and osteogenic lineages [36], but contamination of such cultures from non-myogenic cells is sometimes hard to dismiss as the underlying cause of this observed multipotency [37, 38]. Recent examination using recombination-based lineage tracing indicates that, although satellite cells can be stimulated to accumulate lipid, they do not undergo terminal adipogenic differentiation in vitro [38]. Furthermore, whereas exposure to bone morphogenetic proteins (BMPs) inhibits myogenic differentiation in satellite cells in vitro, it does not result in any overt change to the osteogenic lineage [39]. In vivo, there is also a negligible (<5%) contribution of satellite cells to BMP-mediated ectopic osteogenesis [40]. Therefore, satellite cells can be considered monopotent muscle-resident myogenic stem cells.
Multiple lines of evidence point to functional heterogeneity of satellite cells, which indicates that they do not all have stem cell characteristics. Not only do satellite cell populations from different muscles exhibit heterogeneity in their proliferation rate, clonogenic capacity, extent and rate of differentiation, and ability to self-renew, these variations also exist between satellite cells of the same muscle [37, 41]. Heterogeneity is also revealed by transplantation studies, as only a limited number of grafts result in large numbers of new satellite cells being produced, so extensive self-renewal does not appear to be a universal feature of satellite cells [33, 34].
There are, however, other populations of progenitors isolated from skeletal muscle, including endothelial cells and muscle derived stem cells (MDSCs) [42], blood-vessel associated mesoangioblasts [43], muscle side-population cells [44], CD133+ve cells [45], myoendothelial cells [46] and pericytes [47] (Figure. 5) .
Endothelial cells and muscle derived stem cells MDSCs are multipotent cells, and may involve in skeletal muscle regeneration [20].
Pericytes and mesoangioblasts cells contribute to muscle regeneration after intra-arterial delivery [45].
Since the discovery of (MDSCs), there is convincing evidence that these cells are likely derived from the vascular endothelium and could shares along with satellite cells in regeneration of injuried skeletal muscle [46] (Figure. 6).
MDSCs can be efficiently transduced with antifibrotic and regenerative factors that may enhance skeletal muscle healing [49].
Human (MDSCs) are Characterized by expression of CD13 and to a lesser extent CD10 and CD56, on these cells [50], as well as the lack of hematopoietic marker expression, including CD45. These cells were named mesenchymal stem cells for their ability to differentiate into the mesodermal phenotypes (Figure. 4). For clarity, it should be noted that a population of bone marrow stromal cells has also been termed mesenchymal stem cells, referring to their differentiation potential. Marrow-derived mesenchymal stem cells have been described as expressing the proteins CD29 CD44, CD71, CD90, CD106, CD120a and CD124. It is not clear at this point if these two populations are the same, are distinct, or represent different stages of maturation of the same lineage [51].

11.1. Volumetric Skeletal Muscle Injury
Volumetric muscle loss (VML) is defined as the traumatic or surgical loss of skeletal muscle with resultant functional impairment [26].
11.2. Causes of volumetric muscle loss
Volumetric muscle loss represents a challenging clinical problem for both military and civilian medicine. In military medicine, it occurs primarily from penetrating soft tissue injuries as a result of explosions, and as a consequence of excision and debridement of devitalized muscle tissue following extremity compartment syndrome [53].
In civilian medicine, VML is often associated with penetrating trauma from motor vehicle accidents and sport activities and the implications for the athlete, his/her coach and team may be catastrophic if the injury occurs at a critical time in the athlete's diary. Cranio-maxillo-facial trauma is common on the battlefield [54]. In all cases, VML results in severe cosmetic deformities and debilitating functional loss [26].
11.3. Basic Pathophysiology of Skeletal Muscle Injury
In response to damage, skeletal muscle goes through a well-defined series of events including; degeneration (1to3days), inflammation and regeneration (3 to 4 weeks), fibrosis and extracellular matrix remodeling (3 to 6 months) [9]. The last two phases tend to overlap, (Figure. 7). Ultimately, repair is the result of resident muscle stem cells, known as satellite cells, which proliferate, differentiate, and fuse with existing myofibers or form new myofibers [55]. The normal repair mechanisms, however, are not sufficient for the repair of VML (Kin, 2007).

Mammalian skeletal muscle has an impressive ability to regenerate itself in response to injury [58]. Skeletal muscle repair is a highly synchronized process involving the activation of various cellular and molecular responses, where the coordination between inflammation and regeneration is crucial for the beneficial outcome of the repair process following muscle damage [59].
During muscle tissue repair following damage, the degree of damage and the interactions between muscle and the infiltrating inflammatory cells appear to affect the successful outcome of the muscle repair process. Muscle regeneration depends on a balance between pro-inflammatory and anti-inflammatory factors that determine whether the damage will be resolved with muscle fiber replacement and reconstitution of a functional contractile apparatus, or with scar formation [60]. Although the phases of the repair process are similar after different causes of damage, the kinetics and amplitude of each phase may depend on the particular muscle damaged, the extent of damage, or the damage model used [9].
Immediately following injury, there is a phase of degeneration, which is initiated by the release of proteases into the tissue stroma; these proteases auto digest myofibers and thereby, release tissue debris along the zone of injury [9]. It has been proposed that Tumor necrosis factor-α (TNFα) seems to participate in the muscle protein loss during the degenerative phase of muscle regeneration, and promotes the activation of nuclear factor κB (NFκB) in skeletal muscle cells. NFκB is a transcription factor, which activation alters gene expression and causes proteolysis. In vitro and in vivo data indicate that TNFα promotes the expression of atrogin-1, leading to the catabolism of muscle proteins. This is a result of the activation of the ubiquitin/proteasome pathway in muscle fibers [61, 62].
A rapid necrosis of myofibers is observed. This event is characterized by disruption of the myofiber sarcolemma and results in increased myofiber permeability. As a consequence of the disruption of the myofiber's integrity, the serum levels of muscle proteins, such as creatine kinase, are increased since these proteins are normally restricted to the myofiber cytosol [63]. Within the time frame that this occurs, there is a chemotaxis of neutrophils; the first inflammatory cells to invade the injured muscle. Recent reports suggest that an important increase in their number is observed 1-6 h after muscle damage [64].
About 48 hours after the injury, the primary cells which predominate at the site of injured muscle are macrophages. These cells migrate to the damaged area through the blood stream. Thus, in the case of significant damage of the blood supply to the injured area, regeneration cannot take place until new blood vessels penetrate the area. After the infiltration, macrophages phagocytose cellular debris and remove the disrupted myofilaments, other cytosolic structures and the damaged sarcolemma. Apart from that, macrophages also activate myogenic cells. It appears that the main histopathological characteristics of the early phase of muscle injury are muscle fiber necrosis and an increased number of non-muscles mononucleated cells within the injured area [65].
Although it appears that macrophages may in part be a culprit by initiating inflammatory response, some studies indicate that these cells also secrete various growth factors that directly contribute to tissue regeneration and activation of repair processes necessary for successful recovery from the damage. Additionally, macrophages stimulate the paracrine release of cytokines together with chemotactic factors by T cells, which may locally recruit progenitor and satellite cells towards muscle regeneration [66].
Some of the critical cytokines that orchestrate this local response include, interleukins [IL]-1, -6, and -8 as well as hepatocyte growth factor (HGF), Fibroblast growth factor (FGF) and insulin growth factor [IGF]-1. It is clear from this initial sequence of events that the inflammatory response may be conducive to the repair of skeletal muscle after injury [67].
Interleukin-6 (IL-6) is a cytokine with a major role in the regulation of the inflammation process after muscle injury [68]. IL-6 can be both a factor that induces proteolysis of damaged myofibers and a proliferation signal for satellite cells to replace the destroyed muscle tissue. Some studies suggest that skeletal muscle can produce IL-6 in response to exercise as well as inflammation and injury [69, 70].
Hepatocyte growth factor (HGF) and insulin-like growth factor-1 (IGF-1) are mainly expressed during the regeneration process. Furthermore, histochemical analysis showed that HGF is a protein which is bound to the extracellular matrix in muscle tissue and is released in response to injury. HGF is believed to play a fundamental role during muscle regeneration, especially during the initial phase of muscle repair, and is considered to be essential for effective muscle regeneration [9]. HGF is believed to promote the quiescent satellite cells to enter the cell cycle [29], and expressed mainly during early stages of regeneration, and may, therefore, be available to bind to specific receptors on satellite cells, leading to their activation. It has also been proposed that HGF inhibits the differentiation of myogenic precursor cells [71]. Finally it should be mentioned that HGF could stimulate satellite cells and myogenic precursor cells migration to the site of injury [9].
Fibroblast growth factor (FGF); belongs to a family of cytokines that control cell proliferation, cell differentiation. Several members of the FGF family are expressed in developing skeletal muscle, but FGF-6 is the only one that seems to participate in muscle repair. The high amounts of FGFs, which are released within the short period of inflammation after tissue disruption, may induce satellite cells to proliferate and partly cause the chemotaxis of further muscle precursor cells. Today, it is known that the expression of FGF-6 is stimulated after skeletal muscle injury. FGF-6 induces satellite cell adhesion and compromises their ability to differentiate into myotubes. FGF-6 stimulates the proliferation of satellite cells and induces the expression of genes required for their terminal differentiation [7, 72, and 73].
Insulin growth factor (IGF-1) is important as a survival signal for satellite cells during regeneration and strongly expressed by macrophages and myoblasts during myotube formation and the differentiation process [30, 74]. Moreover, IGF-I promotes muscle cell survival during the initial phase of their differentiation [75].
Equally important is the role of Tumor necrosis factor-α TNFα during the regeneration phase. After skeletal muscle injury TNFα is released not only by infiltrating macrophages, but also by injured muscle fibers [76]. Its expression remains at high levels during the repair process and returns to normal levels several days post-injury [77]. This expression profile, along with in vitro studies, suggests that TNFα has a double role during muscle regeneration: it is both a competence factor (activating satellite cells to enter the cell cycle) and a progression factor (enhancing satellite cell proliferation), once it has been initiated, perhaps via activating the expression of c-fos gene, well known as an important cell growth regulator) [62].
The subsequent phase of myofiber regeneration may begin as early as 24 hours following injury, as evidenced by the cytokine-mediated induction of local satellite cells which previously lie dormant between the basal lamina and sarcolemma. It is not until at least 3 to 5 days after injury, however, that the complete formation of new, centronucleated myofibers can be detected histologically [78].
The characteristics of muscle regeneration are: (a) newly formed fibers are basophilic and this reflects a high protein synthesis. (b) during regeneration, cell fusion is focal to the injured site rather than diffused within the muscle tissue, and (c) the fibers are split, which is probably due to the incomplete fusion of fibers regenerating within the same basal lamina. When fusion of myogenic cells is completed, the size of the newly formed myofibers increases and myonuclei move to the periphery of the fiber. Finally, the new muscle tissue is the same as uninjured muscle, not only morphologically but also functionally [79].
It is likely that a crucial event in the regeneration phase is the differentiation of satellite cells into large numbers of myotubes and myofibers of within just a few days. To date, these progenitor cells are perhaps the best characterized and are often referred to as ‘‘muscle stem cells’’ given their predilection to the myogenic lineage [16].
The progression of activated SCs toward myogenic differentiation followed by fusion into regenerating fibers. The whole process takes approximately 7 days in the mouse while the complete process of regeneration to reform normal tissue architecture typically requires 3 to 4 weeks [80], during which time SCs undergo different fates, giving rise to a few Pax7+ve cells return to quiescence (to maintain the progenitor pool), and many Pax7+ve cells, which are committed to differentiation (Figure. 8) [81].
Perhaps the greatest limitation for patients that results from the pathophysiology of skeletal muscle injury is the formation of dense fibrotic scar tissue. It is clear that fibrosis is induced by a deleterious rise in the cytokine transforming growth factor (TGF)-B1 after injury [82]. The TGF-b1 is a small family of multifunctional growth factor proteins. It has the ability to bind to extracellular proteins and to be stored in the ECM until activation by a physiological process, such as wound healing. After injury, TGF-b1 is released by degranulating platelets at the site of injury and autoinduces its own production by resident cells, such as smooth myogenic cells. Following the denervation and the ischemia of skeletal muscles, TGF-b1 is expressed by regenerating muscles within the first days after a trauma [83].
In the presence of this cytokine, myogenic cells differentiate into myofibroblasts that produce collagen type I, the major component of fibrotic tissue [84]. Ultimately, fibrosis can prevent patients from returning to their baseline function, in part by preventing the formation of new axons toward myofibers and contributes to a decline in muscle contractility and range of motion. The pain that results from fibrosis is also a limiting factor in the recovery of patients during rehabilitation [85].
Although not currently used clinically in this capacity, several agents that block TGFb1 have proven to be remarkably antifibrotic, including gamma-interferon, suramin, and decorin [86, 87]. When suramin was injected immediately or 7 days after the injury, it had only a minor effect on muscle fibrosis. However, when a high dose of suramin was injected 14 days after injury, it prevented fibrosis more effectively than did a lower concentration or no suramin. There were more regenerating myofibers in all the suramin-treated groups than in the control groups. Just as the prevention of fibrosis, the number of regenerating myofibers was dose dependent. Side effects of suramin are adrenocortical insufficiency, malaise, neuropathy, and corneal deposits. Occasionally, neutropenia, thrombocytopenia, and renal failure may occur [88].
Fukushima et al., [89] found that the injection of decorin at 10 and 15 days after injury significantly decreased the amount of fibrosis. Decorin had the additional advantage of enhancing the regeneration of the injured muscle. There seemed to be a dose-response effect.
Therapeutic interventions to improve outcomes following muscle injuries revolve around 3 concepts: enhancing regeneration, blocking fibrosis, and regulating inflammation [57].
Traditional therapies, usually conservative such as non-steroidal anti-inflammatory administration, rest, antioxidant therapy and physical therapy have generally not been helpful in preventing fibrosis and permanent muscle contracture, which leads to functional impairment. Generally, they are not sufficient to enhance muscle regeneration and prevent fibrosis [90].
Extracellular matrix remodeling (ECM); For many years, the ECM was believed to play a passive role during muscle development and regeneration, acting simply as a scaffold for the arrangement of the cells within tissues. Today, it is believed that proteoglycans, collagen, metalloproteinases and other elements of the ECM might play an energetic key role during muscle development [91, 92]. Although the exact function of these other elements in muscle regeneration is not well understood, the similarities between skeletal muscle embryogenesis and regeneration might indicate a similar role of the ECM during these two processes. Satellite cells are surrounded by ECM components and remodeling of these ECM molecules has been seen in many myoblast and muscle cell responses such as migration, fusion and myotube maturation [93].
Moreover, the ECM acts as a reservoir of growth factors. For instance, TGFβ binds to decorin, a heparan sulfate proteoglycan, and is stored in the ECM. Matrix degeneration after injury can release these growth factors, which can then transduce their signals to the cells and regulate the regeneration process [94]. The ECM may also participate in the regulation of the activity of various growth factors. Molecular studies have proven that FGF and TGFβ can bind to extracellular proteoglycans [83]. FGF-binding to syndecan-1 and glypican seems to be essential for the function of FGF during muscle development and repair, while the binding of TGF to decorin inactivates TGF [83, 92]. Therefore, the ECM may be involved in the precise regulation of the activity of growth factors during muscle regeneration
Moreover, it is notable that the ECM possibly participates in satellite cell migration to the site of injury [92, 95]. Cell migration involves a series of complex cell ECM interactions, including adhesion to ECM through integrins, contraction of the cytoskeleton, translocation and release of cell-ECM contact [92]. The right function of integrins is essential for the migration process and may also have a critical role during muscle repair. Furthermore, various studies suggest that interactions between neuron axon and Schwann cells as well as between Schwann cells and the ECM are essential for nerve growth during peripheral nerve regeneration. This suggests a possible pole for ECM in the reinnervation of injured skeletal muscle [96]. Besides, ECM may also participate in signaling pathways essential for muscle differentiation, as myogenin alone cannot stimulate cell terminal differentiation [91].
Another important element of the ECM is the family of matrix metalloproteinases (MMPs) which are a family of Ca- dependent endopeptidases able to cleave most of the ECM components [96]. Their expression is highly regulated during both muscle development and repair [83, 92], and it appears that, in such conditions, MMPs and plasminogen activators act in concert. Thus, plasmin activity together with MMPs is needed to complete a wound healing process; urokinase-type plasminogen activator (uPA) complex participates in fibrinolysis while MMPs have the capacity to split fibrin by acting as pericellular fibrinolysins [92, 97].
To date, data indicate that MMPs are involved in myoblast migration to the site of injury [96]. Urokinase-type plasminogen activator (uPA) and plasmin are implicated in several non-fibrinolytic processes, which lead to ECM degradation, either directly by proteolytic cleavage of ECM components, or indirectly through the activation of latent MMPs [98, 99]. Indeed, a proteolytic activation cascade initiated by uPA/plasmin is involved in MMP activation during muscle regeneration. Plasmin can directly activate several MMPs in vivo through proteolysis and it appears that the activation of MMP-2 and MMP-9 during skeletal muscle regeneration could be mediated by plasmin [99].
Moreover, activated satellite cells can synthesize and secrete MMP-2 and 9 and may be involved in the ECM remodeling after injury, Other MMPs, such as MMP-7, are thought to take part in the fusion of myoblasts [96]. Furthermore, MMPs are possibly involved in the transmission of growth factor signals during muscle repair [83, 92, 96].
11.4. Treatment of volumetric muscle injury
In the 21st century, stem cell therapy has generated a lot of interest in its potential for clinical use in various incurable & chronic diseases [100, 101]. The current standard of care for volumetric muscle injury is to transfer autologous tissue (muscle flaps) using donor tissue from other areas of the victim’s body. Recent reports describe functional free muscle transplantation in the elbow [102], but these procedures were associated with significant donor-site morbidity and are not yet applicable to large defects of load-bearing muscles [103]. The transplantation of stem cells into aberrant or injured tissue has long been a central goal of regenerative medicine and tissue engineering [91].
In addition, nutritional and environmental epigenetics are involved with the repression of anti-aging genes (Sirt 1) that alter gene expression at the cellular level. as a result of changes in cellular chromatin structure, DNA methylation, and histone modifications with relevance to multiple organ dysfunction syndrome [104]. Future research involved with nutritional researches and the maintenance of Sirt 1 transcriptional control is critical to the prevention medicine and correction in epigenetic modifications induced by unhealthy diets or environmental xenobiotics [105] involved in nuclear receptor dysfunction such as the nuclear receptor of anti-aging genes Sirt 1 [106], that determines the survival of man and various species with relevance to toxicity to mitochondria in neurons [107, 108] and cells in peripheral tissues [109-119]. which lead to mitophagy in various tissues [104].
In the developed and developing world nutritional interventions have become essential to prevent Diseases and to maintain the metabolism of glucose, fatty acids, cholesterol, amyloid beta, bile acids and xenobiotics. The World Health Organization has reported that cardiovascular disease is the most prevalent global chronic disease that may be connected to the alarming death rate in various communities. Unhealthy western diets and lifestyles lead to circadian rhythm disorders with defective nutrient and caffeine metabolism associated with cardiovascular disease in the developed world. Caffeine doses in various global populations should be reassessed with relevance to defective anti-aging genes and accelerated mitochondrial apoptosis. Delayed caffeine and xenobiotic metabolism with zinc/magnesium deficiency may be associated to other organ diseases such as gall bladder disease, cardiovascular disease and pancreatic disease in the developing world [120].
Interests in anti-aging therapy with appetite regulation improve an individual’s survival to various chronic diseases [105]. Despite the many studies examining the expression of the anti-aging gene Sirtuin 1 (Sirt 1) with the role of its in the maintenance of other cellular anti-aging genes involved in cell circadian rhythm, senescence, and apoptosis in various models of aging [121], but few studies have focused on the translational effects of SIRT1 expression in muscle to functional relevance [122].
Nowadays, regenerative medicine and tissue engineering for VML therapy is an important treatment for self-renewal and regeneration . while the advancements in skeletal muscle tissue engineering have been remarkable, there are stills several aspects which can be implemented. For example, gene editing with CRISPR/Cas9 can be very beneficial in correcting genetic defects in patient cells thus using the resulting combinations in transplantation procedures. There is no doubt that, once these aspects are improved, the engineering of skeletal muscle tissue using stem cells based therapies will lead to considerable advances in the field of regenerative and personalized medicine [123].
Other researchers have suggested that a biofabrication approach to quickly restore mass of skeletal muscle , functionality and 3D histoarchitecture. By recapitulating muscle anisotropic organization on a microscopic scale, they have been shown to effectively guide cell differentiation and myobundl formation both in vitro and in vivo. Note, during implantation, the biofabricated myosubstitutes favored the formation of new blood vessels and neuromuscular junctions - essential aspects for cell survival and muscle contractile functions - as well as advanced muscle mass and recovery of muscle mass strength. Overall, these strategies represent a solid foundation with further testing and improvements for large animal myosubstitutes, which can be successfully translated into a promising platform in clinical scenarios for reconstructive therapy for patients with VML [124].
It can be concluded that the formation of skeletal muscle begins during the fourth week of embryonic development as specialized mesodermal cells, termed myoblasts, by birth myoblast activity has ceased. Satellite cells are considered to be self-renewing, and serve to generate a population of differentiation-competent myoblasts. Skeletal muscle fibres are classified into three types. The process of contraction, usually triggered by neural impulses, obeys the all-or-none law. VML results in severe cosmetic deformities and debilitating functional loss. Mammalian skeletal muscle has an impressive ability to regenerate itself in response to injury. The transplantation of stem cells into aberrant or injured tissue has long been a central goal of regenerative medicine and tissue engineering.
Clearly Auctoresonline and particularly Psychology and Mental Health Care Journal is dedicated to improving health care services for individuals and populations. The editorial boards' ability to efficiently recognize and share the global importance of health literacy with a variety of stakeholders. Auctoresonline publishing platform can be used to facilitate of optimal client-based services and should be added to health care professionals' repertoire of evidence-based health care resources.

Journal of Clinical Cardiology and Cardiovascular Intervention The submission and review process was adequate. However I think that the publication total value should have been enlightened in early fases. Thank you for all.

Journal of Women Health Care and Issues By the present mail, I want to say thank to you and tour colleagues for facilitating my published article. Specially thank you for the peer review process, support from the editorial office. I appreciate positively the quality of your journal.

Journal of Clinical Research and Reports I would be very delighted to submit my testimonial regarding the reviewer board and the editorial office. The reviewer board were accurate and helpful regarding any modifications for my manuscript. And the editorial office were very helpful and supportive in contacting and monitoring with any update and offering help. It was my pleasure to contribute with your promising Journal and I am looking forward for more collaboration.

We would like to thank the Journal of Thoracic Disease and Cardiothoracic Surgery because of the services they provided us for our articles. The peer-review process was done in a very excellent time manner, and the opinions of the reviewers helped us to improve our manuscript further. The editorial office had an outstanding correspondence with us and guided us in many ways. During a hard time of the pandemic that is affecting every one of us tremendously, the editorial office helped us make everything easier for publishing scientific work. Hope for a more scientific relationship with your Journal.

The peer-review process which consisted high quality queries on the paper. I did answer six reviewers’ questions and comments before the paper was accepted. The support from the editorial office is excellent.

Journal of Neuroscience and Neurological Surgery. I had the experience of publishing a research article recently. The whole process was simple from submission to publication. The reviewers made specific and valuable recommendations and corrections that improved the quality of my publication. I strongly recommend this Journal.

Dr. Katarzyna Byczkowska My testimonial covering: "The peer review process is quick and effective. The support from the editorial office is very professional and friendly. Quality of the Clinical Cardiology and Cardiovascular Interventions is scientific and publishes ground-breaking research on cardiology that is useful for other professionals in the field.

Thank you most sincerely, with regard to the support you have given in relation to the reviewing process and the processing of my article entitled "Large Cell Neuroendocrine Carcinoma of The Prostate Gland: A Review and Update" for publication in your esteemed Journal, Journal of Cancer Research and Cellular Therapeutics". The editorial team has been very supportive.

Testimony of Journal of Clinical Otorhinolaryngology: work with your Reviews has been a educational and constructive experience. The editorial office were very helpful and supportive. It was a pleasure to contribute to your Journal.

Dr. Bernard Terkimbi Utoo, I am happy to publish my scientific work in Journal of Women Health Care and Issues (JWHCI). The manuscript submission was seamless and peer review process was top notch. I was amazed that 4 reviewers worked on the manuscript which made it a highly technical, standard and excellent quality paper. I appreciate the format and consideration for the APC as well as the speed of publication. It is my pleasure to continue with this scientific relationship with the esteem JWHCI.

This is an acknowledgment for peer reviewers, editorial board of Journal of Clinical Research and Reports. They show a lot of consideration for us as publishers for our research article “Evaluation of the different factors associated with side effects of COVID-19 vaccination on medical students, Mutah university, Al-Karak, Jordan”, in a very professional and easy way. This journal is one of outstanding medical journal.

Dear Hao Jiang, to Journal of Nutrition and Food Processing We greatly appreciate the efficient, professional and rapid processing of our paper by your team. If there is anything else we should do, please do not hesitate to let us know. On behalf of my co-authors, we would like to express our great appreciation to editor and reviewers.

As an author who has recently published in the journal "Brain and Neurological Disorders". I am delighted to provide a testimonial on the peer review process, editorial office support, and the overall quality of the journal. The peer review process at Brain and Neurological Disorders is rigorous and meticulous, ensuring that only high-quality, evidence-based research is published. The reviewers are experts in their fields, and their comments and suggestions were constructive and helped improve the quality of my manuscript. The review process was timely and efficient, with clear communication from the editorial office at each stage. The support from the editorial office was exceptional throughout the entire process. The editorial staff was responsive, professional, and always willing to help. They provided valuable guidance on formatting, structure, and ethical considerations, making the submission process seamless. Moreover, they kept me informed about the status of my manuscript and provided timely updates, which made the process less stressful. The journal Brain and Neurological Disorders is of the highest quality, with a strong focus on publishing cutting-edge research in the field of neurology. The articles published in this journal are well-researched, rigorously peer-reviewed, and written by experts in the field. The journal maintains high standards, ensuring that readers are provided with the most up-to-date and reliable information on brain and neurological disorders. In conclusion, I had a wonderful experience publishing in Brain and Neurological Disorders. The peer review process was thorough, the editorial office provided exceptional support, and the journal's quality is second to none. I would highly recommend this journal to any researcher working in the field of neurology and brain disorders.
Dear Agrippa Hilda, Journal of Neuroscience and Neurological Surgery, Editorial Coordinator, I trust this message finds you well. I want to extend my appreciation for considering my article for publication in your esteemed journal. I am pleased to provide a testimonial regarding the peer review process and the support received from your editorial office. The peer review process for my paper was carried out in a highly professional and thorough manner. The feedback and comments provided by the authors were constructive and very useful in improving the quality of the manuscript. This rigorous assessment process undoubtedly contributes to the high standards maintained by your journal.

International Journal of Clinical Case Reports and Reviews. I strongly recommend to consider submitting your work to this high-quality journal. The support and availability of the Editorial staff is outstanding and the review process was both efficient and rigorous.

Thank you very much for publishing my Research Article titled “Comparing Treatment Outcome Of Allergic Rhinitis Patients After Using Fluticasone Nasal Spray And Nasal Douching" in the Journal of Clinical Otorhinolaryngology. As Medical Professionals we are immensely benefited from study of various informative Articles and Papers published in this high quality Journal. I look forward to enriching my knowledge by regular study of the Journal and contribute my future work in the field of ENT through the Journal for use by the medical fraternity. The support from the Editorial office was excellent and very prompt. I also welcome the comments received from the readers of my Research Article.

Dear Erica Kelsey, Editorial Coordinator of Cancer Research and Cellular Therapeutics Our team is very satisfied with the processing of our paper by your journal. That was fast, efficient, rigorous, but without unnecessary complications. We appreciated the very short time between the submission of the paper and its publication on line on your site.

I am very glad to say that the peer review process is very successful and fast and support from the Editorial Office. Therefore, I would like to continue our scientific relationship for a long time. And I especially thank you for your kindly attention towards my article. Have a good day!

"We recently published an article entitled “Influence of beta-Cyclodextrins upon the Degradation of Carbofuran Derivatives under Alkaline Conditions" in the Journal of “Pesticides and Biofertilizers” to show that the cyclodextrins protect the carbamates increasing their half-life time in the presence of basic conditions This will be very helpful to understand carbofuran behaviour in the analytical, agro-environmental and food areas. We greatly appreciated the interaction with the editor and the editorial team; we were particularly well accompanied during the course of the revision process, since all various steps towards publication were short and without delay".

I would like to express my gratitude towards you process of article review and submission. I found this to be very fair and expedient. Your follow up has been excellent. I have many publications in national and international journal and your process has been one of the best so far. Keep up the great work.

We are grateful for this opportunity to provide a glowing recommendation to the Journal of Psychiatry and Psychotherapy. We found that the editorial team were very supportive, helpful, kept us abreast of timelines and over all very professional in nature. The peer review process was rigorous, efficient and constructive that really enhanced our article submission. The experience with this journal remains one of our best ever and we look forward to providing future submissions in the near future.

I am very pleased to serve as EBM of the journal, I hope many years of my experience in stem cells can help the journal from one way or another. As we know, stem cells hold great potential for regenerative medicine, which are mostly used to promote the repair response of diseased, dysfunctional or injured tissue using stem cells or their derivatives. I think Stem Cell Research and Therapeutics International is a great platform to publish and share the understanding towards the biology and translational or clinical application of stem cells.

I would like to give my testimony in the support I have got by the peer review process and to support the editorial office where they were of asset to support young author like me to be encouraged to publish their work in your respected journal and globalize and share knowledge across the globe. I really give my great gratitude to your journal and the peer review including the editorial office.

I am delighted to publish our manuscript entitled "A Perspective on Cocaine Induced Stroke - Its Mechanisms and Management" in the Journal of Neuroscience and Neurological Surgery. The peer review process, support from the editorial office, and quality of the journal are excellent. The manuscripts published are of high quality and of excellent scientific value. I recommend this journal very much to colleagues.

Dr.Tania Muñoz, My experience as researcher and author of a review article in The Journal Clinical Cardiology and Interventions has been very enriching and stimulating. The editorial team is excellent, performs its work with absolute responsibility and delivery. They are proactive, dynamic and receptive to all proposals. Supporting at all times the vast universe of authors who choose them as an option for publication. The team of review specialists, members of the editorial board, are brilliant professionals, with remarkable performance in medical research and scientific methodology. Together they form a frontline team that consolidates the JCCI as a magnificent option for the publication and review of high-level medical articles and broad collective interest. I am honored to be able to share my review article and open to receive all your comments.

“The peer review process of JPMHC is quick and effective. Authors are benefited by good and professional reviewers with huge experience in the field of psychology and mental health. The support from the editorial office is very professional. People to contact to are friendly and happy to help and assist any query authors might have. Quality of the Journal is scientific and publishes ground-breaking research on mental health that is useful for other professionals in the field”.

Dear editorial department: On behalf of our team, I hereby certify the reliability and superiority of the International Journal of Clinical Case Reports and Reviews in the peer review process, editorial support, and journal quality. Firstly, the peer review process of the International Journal of Clinical Case Reports and Reviews is rigorous, fair, transparent, fast, and of high quality. The editorial department invites experts from relevant fields as anonymous reviewers to review all submitted manuscripts. These experts have rich academic backgrounds and experience, and can accurately evaluate the academic quality, originality, and suitability of manuscripts. The editorial department is committed to ensuring the rigor of the peer review process, while also making every effort to ensure a fast review cycle to meet the needs of authors and the academic community. Secondly, the editorial team of the International Journal of Clinical Case Reports and Reviews is composed of a group of senior scholars and professionals with rich experience and professional knowledge in related fields. The editorial department is committed to assisting authors in improving their manuscripts, ensuring their academic accuracy, clarity, and completeness. Editors actively collaborate with authors, providing useful suggestions and feedback to promote the improvement and development of the manuscript. We believe that the support of the editorial department is one of the key factors in ensuring the quality of the journal. Finally, the International Journal of Clinical Case Reports and Reviews is renowned for its high- quality articles and strict academic standards. The editorial department is committed to publishing innovative and academically valuable research results to promote the development and progress of related fields. The International Journal of Clinical Case Reports and Reviews is reasonably priced and ensures excellent service and quality ratio, allowing authors to obtain high-level academic publishing opportunities in an affordable manner. I hereby solemnly declare that the International Journal of Clinical Case Reports and Reviews has a high level of credibility and superiority in terms of peer review process, editorial support, reasonable fees, and journal quality. Sincerely, Rui Tao.

Clinical Cardiology and Cardiovascular Interventions I testity the covering of the peer review process, support from the editorial office, and quality of the journal.
